
Flail (weapon)

A flail is a weapon consisting of a striking head attached to a handle by a flexible rope, strap, or chain. The chief tactical virtue of the flail was its capacity to strike around a defender's shield or parry. Its chief liability was a lack of precision and the difficulty of using it in close combat, or closely-ranked formations.
There are two broad types of flail: a long, two-handed infantry weapon with a cylindrical head, and a shorter weapon with a round metal striking head. The longer cylindrical-headed flail is a hand weapon derived from the agricultural tool of the same name, commonly used in threshing. It was primarily considered a peasant's weapon, and while not common, they were deployed in Germany and Central Europe in the later Late Middle Ages.[1] The smaller, more spherical-headed flail appears to be even less common; it appears occasionally in artwork from the 15th century onward, but many historians have expressed doubts that it ever saw use as an actual military weapon.
The peasant flail
[edit]
In the Late Middle Ages, a particular type of flail appears in several works being used as a weapon, which consists of a very long shaft with a hinged, roughly cylindrical striking end. In most cases, these are two-handed agricultural flails, which were sometimes employed as an improvised weapon by peasant armies conscripted into military service or engaged in popular uprisings. For example, in the 1420–1497 period, the Hussites fielded large numbers of peasant foot soldiers armed with this type of flail.[1][2][3]
Some of these weapons featured anti personnel studs or spikes embedded in the striking end, or are shown being used by armored knights,[4] suggesting they were made or at least modified specifically to be used as weapons. Such modified flails were used in the German Peasants' War in the early 16th century.[5][6] Several German martial arts manuals or Fechtbücher from the 15th, 16th and 17th century feature illustrations and lessons on how to use the peasant flail (with or without spikes) or how to defend against it when attacked.[7][8][9][10]
The military flail
[edit]


The other type of European flail is a shorter weapon consisting of a wooden haft connected by a chain, rope, or leather to one or more striking ends. The kisten, with a spiked or non-spiked head and a leather or rope connection to the haft, is attested in the 10th century in the territories of the Rus', probably being adopted from either the Avars or Khazars. This weapon spread into Central and Eastern Europe in the 11th–13th centuries,[11] and then further west in Western Europe during the 12th and 13th centuries.[12] The medieval military flail (fléau d'armes in French and Kriegsflegel in German), then, might typically have consisted of a wooden shaft joined by a length of chain to one or more iron-shod wooden bars,[13] or it may have been a Kettenmorgenstern ("chain morning star") with one or more metal balls or morning star in the place of the wooden bars.[14] Artwork from the 15th century to the early 17th century shows most of these weapons having handles longer than 3 ft and being wielded with two hands, but a few are shown used in a single hand or with a haft too short to be used two-handed.
Despite being very common in fictional works such as cartoons, films and role-playing games as a "quintessential medieval weapon", historical information about flails other than the kisten or derivatives of the peasant flail is rarer than other contemporary weapons, but a notable body of visual and textual sources for Western, Central, and Southern European depictions and descriptions of military are extant, if not particularly easy to find.[12] Some doubt they were used as weapons at all due to the scarcity of genuine specimens as well as the unrealistic way they are depicted in art, as well as the number of pieces in museums that turned out to be 19th century forgeries when analyzed,[15][16][17] though these limited and somewhat sensationlist studies have now been largely debunked.[12] Waldman (2005) documented several likely authentic examples of the ball-and-chain flail from private collections as well as several restored illustrations from German, French, and Czech sources.[18] Even the more comprehensive scholarly articles collating the numerous sources for flails note that their use in warfare was likely rare at best, even if such weapons were known about as a concept.[12] Flails are noted as being potentially hazardous to their user in the absence of appropriate training an experience,[16] meaning that, even if a blow were struck, there may have been a long time before the user could ready another swing.[18]
Variations outside Europe
[edit]In Asia, short flails originally employed in threshing rice were adapted into weapons such as the nunchaku or three-section staff. In China, a very similar weapon to the long-handled peasant flail is known as the two-section staff, and Korea has a weapon called a pyeongon.[19][20][21] In Japan, there is also a version of the smaller ball-on-a-chain flail called a chigiriki.
In the 18th and 19th centuries, the long-handled flail is found in use in India. An example held in the Pitt Rivers Museum has a wooden ball-shaped head studded with iron spikes. Another in the Royal Armouries collection has two spiked iron balls attached by separate chains.
The knout, a whip or scourge formerly used in Russia for the punishment of criminals, was the descendant of the flail. It was manufactured in many forms, and its effect was so severe that few of those who were subjected to its full force survived the punishment. The Emperor Nicholas I substituted a milder whip for the knout.[22]
Gallery
[edit]-
 a representative Ottoman flail (replica)
a representative Ottoman flail (replica) -
 Hayo van Wolvega attacks the tower of Damietta with a flail during the 5th crusade
Hayo van Wolvega attacks the tower of Damietta with a flail during the 5th crusade -
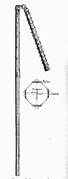 A two-handed flail with metal studs
A two-handed flail with metal studs -
 Detail from The Travels of Marco Polo, circa 1410, showing an armored "Mamluk" with a short, spiked flail tucked into his belt
Detail from The Travels of Marco Polo, circa 1410, showing an armored "Mamluk" with a short, spiked flail tucked into his belt -
 Detail from The Travels of Marco Polo, circa 1410, showing a horseman using a spiked flail with both hands to strike an adversary.
Detail from The Travels of Marco Polo, circa 1410, showing a horseman using a spiked flail with both hands to strike an adversary. -
 Illustration from Bellifortis showing a mounted knight with a short flail, circa 1450.
Illustration from Bellifortis showing a mounted knight with a short flail, circa 1450. -
 Hussite troops with flails on the march
Hussite troops with flails on the march







See also
[edit]References
[edit]- ^ a b Eduard Wagner; Zoroslava Drobná; Jan Durdík (5 May 2014). Medieval Costume, Armour and Weapons. Courier Corporation. ISBN 978-0-486-32025-0.
- ^ Stephen Turnbull: The Hussite Wars 1419-36, Osprey MAA 409,2004
- ^ media:344Wagenburg der Hussiten.jpg media:Hussites massacre.jpg
- ^ Maximilian I. "Colored plate depicting knights fighting with two-handed flails". Freydal. Archived from the original on 2007-07-02. Retrieved 2016-01-19.
- ^ Douglas Miller : Armies of the German Peasant's War 1524-26,Osprey MAA 384,2003
- ^ media:German Peasants War.jpg
- ^ Hans Talhoffer (c. 1450s). "Talhoffer Fechtbuch (MS 78.A.15) Folio 60r". wiktenauer.com. Retrieved 2016-02-01.
- ^ Hans Talhoffer (c. 1450s). "Talhoffer Fechtbuch (MS 78.A.15) Folio 60v". wiktenauer.com. Retrieved 2016-02-01.
- ^ Michael Hundt (1611). "Ein new Kůnstliches Fechtbuch im Rappier - Figure 88". wiktenauer.com. Retrieved 2016-02-01.
- ^ Jakob Sutor von Baden (1612). "New Kůnstliches Fechtbuch - Page 108". wiktenauer.com. Retrieved 2016-02-01.
- ^ Kotowicz, Piotr N. (2008). "Early medieval war-flails (kistens) from Polish lands" (PDF). Fasciculi Archaeologiae Historicae. XXI: 75–86.
- ^ a b c d Holdsworth, Alistair F. (March 2024). "Fantastic Flails and Where to Find Them: The Body of Evidence for the Existence of Flails in the Early and High Medieval Eras in Western, Central, and Southern Europe". Histories. 4 (1): 144–203. doi:10.3390/histories4010009. ISSN 2409-9252.
- ^ Cowper, Henry Swainson (1906). The art of attack: Being a study in the development of weapons and appliances of offence, from the earliest times to the age of gunpowder. Ulverston: W. Holmes, ltd., Printers. p. https://archive.org/details/artattack00cowpgoog/page/n106 80].
- ^ DeVries, Kelly (2012). Medieval military technology. North York, Ont. Tonawanda, N.Y: University of Toronto Press. p. 30. ISBN 978-1-4426-0497-1.
- ^ Dr. Paul B. Sturtevant (May 12, 2016). "The Curious Case of the Weapon that Didn't Exist". The Public Medievalist. Retrieved 2016-06-01.
- ^ a b DeVries, Kelly (2007). Medieval weapons an illustrated history of their impact. Santa Barbara, Calif: ABC-CLIO. p. 133. ISBN 978-1-85109-526-1.
- ^ "Military Flail" catalog descriptions ([1], [2], [3]; see especially "Date" field) at the Metropolitan Museum of Art. Accessed 2015-02-24.
- ^ a b Waldman, John (2005). Hafted weapons in medieval and Renaissance Europe the evolution of European staff weapons between 1200 and 1650. Boston: Brill. pp. 145–150. ISBN 90-04-14409-9.
- ^ "네이버 지식iN :: 지식과 내가 함께 커가는 곳". Kin.naver.com. Retrieved 2012-12-18.
- ^ "네이버 지식백과". 100.naver.com. Retrieved 2012-12-18.
- ^ "VOTE!". Dvdprime.donga.com. 2009-05-08. Archived from the original on 2014-01-18. Retrieved 2012-12-18.
- ^ Sargeaunt, Bertram Edward (1908). Weapons, a brief discourse on hand-weapons other than fire-arms. London, H. Rees. p. 11.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}